






122-123 


Метаболізм. регуляція
гормональний контроль
 Каталізуються ферментами активація і відповідно інактивація ключових ферментів проміжного метаболізму називаються взаємоперетворення. Такі процеси перебувають під різноманітним контролем, і тому числі і гормональних. У цьому розділі розглянуто процеси взаємоперетворення, які здійснюють регуляцію метаболізму глікогену в печінці.
Каталізуються ферментами активація і відповідно інактивація ключових ферментів проміжного метаболізму називаються взаємоперетворення. Такі процеси перебувають під різноманітним контролем, і тому числі і гормональних. У цьому розділі розглянуто процеси взаємоперетворення, які здійснюють регуляцію метаболізму глікогену в печінці.
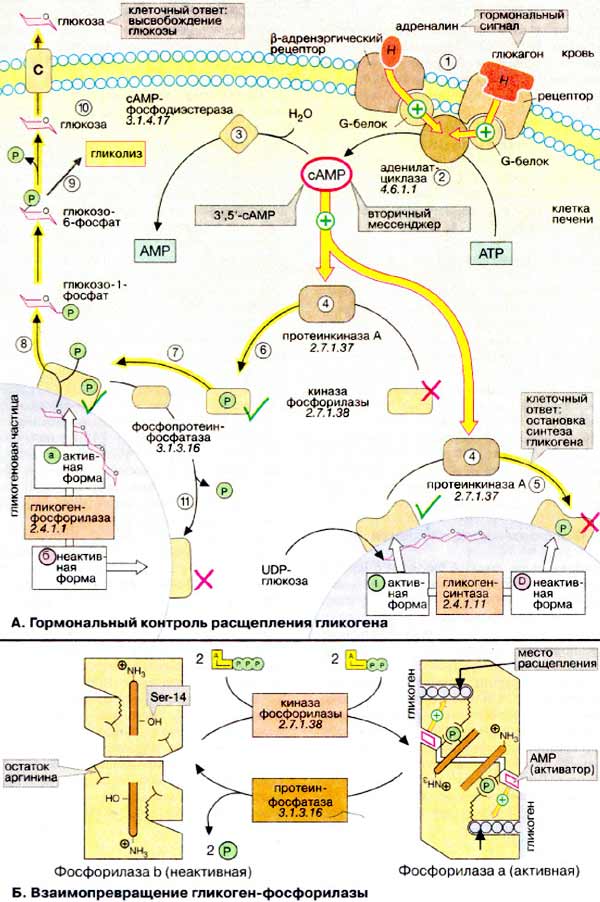
А. Гормональний контроль розщеплення глікогену
Глікоген служить в організмі резервом вуглеводів, з якого в печінці і м'язах шляхом розщеплення швидко створюється глюкозофосфат (див. С. 158 ). Швидкість синтезу глікогену визначається активністю глікоген-синтази (на схемі внизу праворуч), в той час як розщеплення каталізується глікоген-фосфорілазой (на схемі внизу зліва). Обидва ферменти діють на поверхні нерозчинних частинок глікогену, де вони в залежності від стану обміну речовин можуть перебувати в активній або неактивній формі. При голодуванні або в стресових ситуаціях (боротьба, біг) зростає потреба організму в глюкозі. У таких випадках виділяються гормони адреналін і глюкагон. Вони активують розщеплення і пригнічують синтез глікогену. Адреналін діє в м'язах і печінці, а глюкагон - тільки в печінці.
Обидва гормону зв'язуються з рецепторами на плазматичній мембрані (1) і активують за посередництва G-білків (див. С. 372 ) Аденилатциклазу (2), яка каталізує синтез 3 ', 5'-цикло-AMФ (цАМФ) з АТФ (АТР). Дзеркально протилежним є дія на цей 'вторинний месенджер' фосфодіестерази цАМФ (3), гідроліз цАМФ до АМФ (AMP). У печінки діестераза індукується інсуліном, який тому не перешкоджає впливу двох інших гормонів (не показано). цАМФ зв'язується і тим самим активує протеїн А (4), яка діє за двома напрямками: з одного боку, за допомогою фосфорилювання за участю АТФ в якості коферменту вона переводить в неактивну D-форму глікоген-синтази і внаслідок цього зупиняє синтез глікогену (5) ; з іншого, активує - також шляхомфосфорилювання - іншу протеинкиназу, киназу фосфорілази (8). Активна киназа фосфорілази фосфорилирует неактивну b-форму глікоген-фосфорилази, перетворюючи її в активну а-форму (7). Це призводить до вивільнення з глікогену глюкозо-1-фосфату (8), який після перетворення в глюкозо-6-фосфат з участю фосфоглюкомутази включається в гліколіз (9). У печінці додатково утворюється вільна глюкоза, яка надходить у кров (10).
У міру зменшення рівня цАМФ активуються фосфопротеінфосфатази (11), які дефосфорилюється різні Фосфопротеіни описаного каскаду і тим самим зупиняють розщеплення глікогену і ініціюють його синтез. Ці процеси протікають протягом декількох секунд, так що метаболізм глікогену швидко адаптується до змінених умов.
Б. Взаємоперетворення глікоген-фосфорилази
Структурні зміни, які супроводжують взаємоперетворення глікоген-фосфорилази, були встановлені рентгеноструктурньїм аналізом. Фермент являє собою димер з симетрією другого порядку. Кожна субодиниця має активний центр, який розташований всередині білка і в b-формі погано доступний для субстрату. Взаємоперетворенням починається з фосфорилювання серинового залишку (Ser-14) поблизу N-кінця кожної з субодиниць. З фосфатними групами зв'язуються залишки аргініну сусідніх субодиниць. Зв'язування ініціює конформаційні перебудови, які суттєво збільшують спорідненість ферменту до аллостеріческому активатора АМФ. Дія АМФ і вплив конформаційних змін на активні центри призводять до виникнення більш активної а-форми. Після видалення фосфатних залишків фермент мимовільно приймає вихідну b-конформацію.