






Сюрпризи мітохондріального геному
Г.М. Димшиц
Григорій Мойсейович Димшиц,
доктор біологічних наук, професор кафедри молекулярної біології Новосибірського державного університету, завідувач лабораторією структури генома Інституту цитології і генетики Сибірського відділення РАН. Співавтор і редактор чотирьох шкільних підручників із загальної біології. З часу виявлення в мітохондріях молекул ДНК минуло чверть століття, перш ніж ними зацікавилися не лише молекулярні біологи і цитологи, але і генетики, еволюціоністи, а також палеонтологи і криміналісти, історики та лінгвісти. Такий широкий інтерес спровокувала робота А.Уілсона з Каліфорнійського університету. У 1987 р він опублікував результати порівняльного аналізу ДНК мітохондрій, взятих у 147 представників різних етносів всіх людських рас, що заселяють п'ять континентів. За типом, місцем розташування та кількості індивідуальних мутацій встановили, що всі мітохондріальні ДНК виникли з однієї предковой послідовності нуклеотидів шляхом дивергенції. У околонаучной пресі висновок цей інтерпретували вкрай спрощено - все людство походить від однієї жінки, названої мітохондріальної Євою (і дочки і сини отримують мітохондрії тільки від матері), яка жила в Північно-Східній Африці близько 200 тис. Років тому. Ще через 10 років вдалося розшифрувати фрагмент ДНК мітохондрій, виділений з останків неандертальця, і оцінити час існування останнього загального предка людини і неандертальця в 500 тис. Років тому [ 1 ].
Сьогодні мітохондріальна генетика людини інтенсивно розвивається як в популяційному, так і в медичному аспекті. Встановлено зв'язок між рядом важких спадкових захворювань і дефектами в мітохондріальних ДНК. Генетичні зміни, асоційовані зі старінням організму, найбільш виражені в мітохондріях. Що ж представляє із себе геном мітохондрій, що відрізняється у людини і інших тварин від такого у рослин, грибів і найпростіших і за розміром, і за формою, і за генетичною ємності? Як працює і як виник мітохондріальний геном у різних таксонів? Про це і піде мова в нашій статті.
У всіх евкаріот - будь це малярійний плазмодій, дрібний одноклітинний паразит, що руйнує еритроцити людини, або сама людина, гігантська свободноживущими клітина амеба протей, мікроскопічна колонія дріжджів або гриб, який має багатокілометровий міцелій, ефемерні комахи поденки або тисячолітні секвої - у всіх генетична інформація міститься не тільки в хромосомах клітинного ядра, але і в мітохондріях - самовідтворюються напівавтономних органелах клітини, що мають власний геном. У той час як ядерний геном являє собою сукупність лінійних молекул ДНК гаплоїдного набору хромосом, мітохондріальний геном - одну або кілька кільцевих (рідко лінійних) молекул ДНК (мтДНК). У виняткових випадках евкаріотіческіе клітини не містять мітохондрій, наприклад деякі паразитуючі в кишечнику анаеробні амеби.
Мітохондрії називають енергетичними станціями клітини. Крім зовнішньої гладкою мембрани вони мають внутрішню мембрану, що утворить численні складки - Крісті. У них вбудовані білкові компоненти дихального ланцюга - ферменти, що беруть участь в перетворенні енергії хімічних зв'язків окислюваних поживних речовин в енергію молекул аденозинтрифосфорної кислоти (АТФ). Такий "конвертованою валютою" клітина оплачує всі свої енергетичні потреби. У клітинах зелених рослин крім мітохондрій є ще й інші енергетичні станції - хлоропласти. Вони працюють на "сонячних батареях", але теж утворюють АТФ з АДФ і фосфату. Як і мітохондрії, хлоропласти - автономно розмножуються органели - також мають дві мембрани і містять ДНК.
У матриксі мітохондрій, крім ДНК, знаходяться і власні рибосоми, за багатьма характеристиками відрізняються від евкаріотіческіх рибосом, розташованих на мембранах ендоплазматичної мережі. Однак на рибосомах мітохондрій утворюється не більше 5% від всіх білків, що входять до їх складу. Більша частина білків, складових структурні і функціональні компоненти мітохондрій, кодується ядерним геномом, синтезується на рибосомах ендоплазматичної мережі і транспортується по її каналах до місця збірки. Таким чином, мітохондрії - це результат об'єднаних зусиль двох геномів і двох апаратів транскрипції і трансляції. Деякі суб'едінічние ферменти дихального ланцюга мітохондрій складаються з різних поліпептидів, частина яких кодується ядерним, а частина - мітохондріальних геномом. Наприклад, ключовий фермент окисного фосфорилювання - цитохром-с-оксидаза у дріжджів складається з трьох субодиниць, кодованих і синтезованих в мітохондріях, і чотирьох, кодованих в ядрі клітини і синтезованих в цитоплазмі. Експресією більшості генів мітохондрій управляють певні гени ядер.
Розміри і форми мітохондріальних геномів
До теперішнього часу прочитано більше 100 різних геномів мітохондрій. Набір і кількість їх генів в мітохондріальних ДНК, для яких повністю визначена послідовність нуклеотидів, сильно розрізняються у різних видів тварин, рослин, грибів і найпростіших. Найбільша кількість генів виявлено в мітохондріальному геномі жгутикового найпростішого Rectinomonas americana - 97 генів, включаючи всі кодують білок гени, знайдені в мтДНК інших організмів. У більшості вищих тварин геном мітохондрій містить 37 генів: 13 для білків дихального ланцюга, 22 для тРНК і два для рРНК (для великої субодиниці рибосом 16S рРНК і для малої 12S рРНК). У рослин і найпростіших, на відміну від тварин і більшості грибів, в мітохондріальному геномі закодовані і деякі білки, що входять до складу рибосом цих органел. Ключові ферменти матричного полінуклеотидні синтезу, такі як ДНК-полімераза (здійснює реплікацію мітохондріальної ДНК) і РНК-полімераза (транскрибується геном мітохондрій), зашифровані в ядрі і синтезуються на рибосомах цитоплазми. Цей факт вказує на відносність автономії мітохондрій у складній ієрархії евкаріотіческой клітини.
Геноми мітохондрій різних видів відрізняються не тільки по набору генів, порядку їх розташування та експресії, але за розміром і формою ДНК. Переважна більшість описаних сьогодні мітохондріальних геномів є кільцеві суперспіралізованние двуцепочечние молекули ДНК. У деяких рослин поряд з кільцевими формами є і лінійні, а у деяких найпростіших, наприклад інфузорій, в мітохондріях виявлені тільки лінійні ДНК [ 2 ].
Як правило, в кожній мітохондрії міститься кілька копій її генома. Так, в клітинах печінки людини близько 2 тис. Мітохондрій, і в кожній з них - по 10 однакових геномів. У фібробластах миші 500 мітохондрій, що містять по два генома, а в клітинах дріжджів S.cerevisiae - до 22 мітохондрій, що мають по чотири геному.
Мітохондріальний геном рослин, як правило, складається з декількох молекул різного розміру. Одна з них, "основна хромосома", містить велику частину генів, а кільцеві форми меншої довжини, що знаходяться в динамічній рівновазі як між собою, так і з основною хромосомою, утворюються в результаті внутрішньо-і міжмолекулярної рекомбінації завдяки наявності повторених послідовностей (рис.1 ).

Рис 1. Схема освіти кільцевих молекул ДНК різного розміру в мітохондріях рослин.
Рекомбінація відбувається по повторенням ділянок (позначені синім кольором).
В мітохондріях більшості організмів (крім вищих тварин) частину кільцевих молекул ДНК присутня у вигляді олігомерів, які можна розділити на три класи: лінійні; кільцеві, що мають контурну довжину, кратну довжині мономірних кілець; ланцюгові, Катена, що складаються з топологічно пов'язаних, тобто просмикнутих один в одного, мономірних кілець (рис.2). Так, в єдиній мітохондрії найпростіших із загону кінетопластиди, що включає Ендопаразити людини - трипаносом, містяться тисячі кільцевих молекул ДНК. У Trypanosoma brucei є два типи молекул: 45 однакових максі-кілець, кожне з яких складається з 21 тис. Пар нуклеотидів, і 5.5 тис. Ідентичних один одному міні-кілець по 1000 пар нуклеотидів. Всі вони, з'єднуючись в Катена, утворюють переплетену мережу, яка разом з білками формує структуру, яка називається кінетопласт.
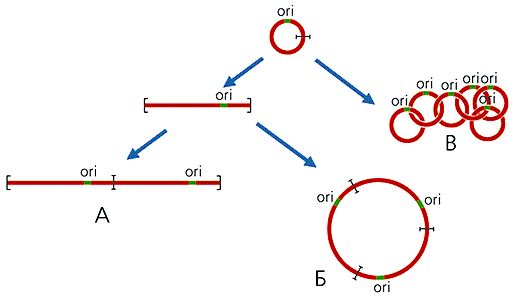
Рис 2. Схема освіти лінійних (А), кільцевих (Б), ланцюгових (В) олігомерів мтДНК.
ori - район початку реплікації ДНК.
Розмір геному мітохондрій різних організмів коливається від менш 6 тис. Пар нуклеотидів у плазмодія (в ньому, крім двох генів рРНК, міститься тільки три гени, що кодують білки) до сотень тисяч пар нуклеотидів у наземних рослин (наприклад, у Arabidopsis thaliana з сімейства хрестоцвітних 366924 пар нуклеотидів). При цьому 7-8-кратні відмінності в розмірах мтДНК вищих рослин виявляються навіть в межах одного сімейства. Довжина мтДНК хребетних тварин відрізняється незначно: у людини - 16569 пар нуклеотидів, у свині - 16350, у дельфіна - 16330, у шпорцевой жаби Xenopus laevis - 17533, у коропа - 16400. Ці геноми подібні також і по локалізації генів, більшість яких розташовуються встик ; в ряді випадків вони навіть перекриваються, зазвичай на один нуклеотид, так що останній нуклеотид одного гена виявляється першим в наступному. На відміну від хребетних, у рослин, грибів і найпростіших мтДНК містять до 80% не кодують послідовностей. У різних видів порядок генів в геномах мітохондрій відрізняється.
Висока концентрація активних форм кисню в мітохондріях і слабка система репарації збільшують частоту мутацій мтДНК у порівнянні з ядерної на порядок. Радикали кисню служать причиною специфічних замін Ц ® Т (дезамінування цитозину) і Г ® Т (окисне пошкодження гуаніну), внаслідок чого, можливо, мтДНК багаті АТ-парами. Крім того, всі мтДНК мають цікавим властивістю - вони не метіліруется, на відміну від ядерних і прокариотических ДНК. Відомо, що метилування (тимчасова хімічна модифікація нуклеотидноїпослідовності без порушення кодує функції ДНК) - один з механізмів програмованої інактивації генів [ 3 ].
Реплікація і транскрипція ДНК мітохондрій ссавців
У більшості тварин комплементарні ланцюга в мтДНК значно розрізняються по питомій щільності, оскільки містять неоднакову кількість "важких" пуринових і "легких" піримідинових нуклеотидів. Так вони і називаються - H (heavy - важка) і L (light - легка) ланцюг. На початку реплікації молекули мтДНК утворюється так звана D-петля (від англ. Displacement loop - петля зміщення). Ця структура, видима в електронний мікроскоп, складається з двуцепочечной і одноланцюжкові (відсунутою частини Н-ланцюги) ділянок. Двуцепочечной ділянку формується частиною L-ланцюга і комплементарних їй знову синтезованим фрагментом ДНК довжиною 450-650 (в залежності від виду організму) нуклеотидів, які мають на 5'-кінці рібонуклеотідную приманку, яка відповідає точці початку синтезу Н-ланцюги (ori H). Синтез L-ланцюга починається лише тоді, коли дочірня Н-ланцюг доходить до точки ori L. Це обумовлено тим, що область ініціації реплікації L-ланцюга доступна для ферментів синтезу ДНК лише в одноланцюговий стані, а отже, тільки в розплетеною подвійної спіралі при синтезі Н-ланцюги. Таким чином, дочірні ланцюга мтДНК синтезуються безперервно і асинхронно (рис.3).

Рис 3. Схема реплікації мтДНК ссавців.
Спочатку формується D-петля, потім синтезується дочірня Н-ланцюг,
потім починається синтез дочірньої L-ланцюга.
В мітохондріях загальне число молекул з D-петлею значно перевищує число повністю реплицирующихся молекул. Обумовлено це тим, що у D-петлі є додаткові функції - прикріплення мтДНК до внутрішньої мембрані і ініціація транскрипції, оскільки в цьому районі локалізовано промотори транскрипції обох ланцюгів ДНК.
На відміну від більшості евкаріотіческіх генів, які транскрибуються незалежно один від одного, кожна з ланцюгів мтДНК ссавців переписується з утворенням однієї молекули РНК, що починається в районі ori H. Крім цих двох довгих молекул РНК, комплементарних Н і L-ланцюгів, формуються і більш короткі ділянки Н-ланцюги, які починаються в тій же точці і закінчуються на 3'-кінці гена 16S рРНК (рис.4). Таких коротких транскриптов в 10 разів більше, ніж довгих. В результаті дозрівання (процесингу) з них утворюються 12S рРНК і 16S рРНК, які беруть участь у формуванні мітохондріальних рибосом, а також фенілаланіновой і валіновая тРНК. З довгих транскриптов вирізаються інші тРНК і утворюються транслюються мРНК, до 3'-кінців яких приєднуються поліаденіловой послідовності. 5'-кінці цих мРНК НЕ кепіруются, що незвично для евкаріот. Сплайсингу (зрощування) не відбувається, оскільки жоден з мітохондріальних генів ссавців не містить інтронів.

ND1-ND6, ND4L - гени субодиниць нaд-H-дегідрогеназну комплексу;
СОI-COIII - гени субодиниць цитохром-с-оксидази;
ATP6, ATP8 - гени субодиниць AТФ-синтетази
Cyt b - ген цитохрома b.Сюрпризи мітохондріального геному
Незважаючи на те, що в геномах мітохондрій ссавців і дріжджів міститься приблизно однакова кількість генів, розміри дріжджового геному в 4-5 разів більше - близько 80 тис. Пар нуклеотидів. Хоча кодують послідовності мтДНК дріжджів високо гомологічних відповідним послідовностям у людини, дріжджові мРНК додатково мають 5'-лідерних і 3'-Некодуючі області, як і більшість ядерних мРНК. Ряд генів містить ще й інтрони. Так, в гені box, що кодує цитохромоксидазу b, є два інтрони. З первинного РНК-транскрипту автокаталитически (без участі будь-яких білків) вирізається копія більшої частини першого інтрони. Частина, що залишилася РНК служить матрицею для утворення ферменту матурази, що бере участь в сплайсинге. Частина її амінокислотноїпослідовності закодована в останніх копіях интронов. Матураза вирізає їх, руйнуючи свою власну мРНК, копії екзонів зшиваються, і утворюється мРНК для цитохромоксидази b (рис.5). Відкриття такого феномена змусило переглянути уявлення про інтрони, як про "нічого не кодують послідовностях".
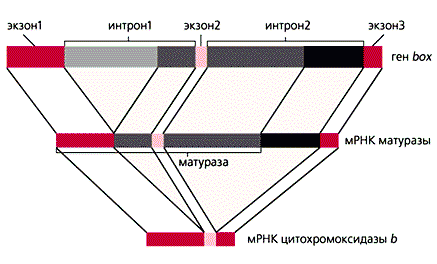
Рис 5. Процессинг (дозрівання) мРНК цитохромоксидази b в мітохондріях дріжджів.
На першому етапі сплайсингу утворюється мРНК, по якій синтезується матураза,
необхідна для другого етапу сплайсингу.
При вивченні експресії мітохондріальних генів Trypanosoma brucei виявилося дивовижне відхилення від однієї з основних аксіом молекулярної біології, яка говорить, що послідовність нуклеотидів в мРНК в точності відповідає такій в кодують ділянках ДНК. Виявилося, мРНК однією з субодиниць цитохром-с-оксидази редагується, тобто після транскрипції змінюється її первинна структура - вставляється чотири урацила. В результаті утворюється нова мРНК, що служить матрицею для синтезу додаткової субодиниці ферменту, послідовність амінокислот в якій не має нічого спільного з послідовністю, кодируемой нередактірованной мРНК (див. Таблицю).

Відбувається це за рахунок зсуву рамки зчитування на число нуклеотидів, що не кратне розміру триплета (в даному випадку на чотири). Нова білкова субодиниця, необхідна для роботи ферменту, утворюється в мітохондріях паразита тільки тоді, коли він потрапляє в організм холоднокровних мухи і потребує окисного фосфорилювання для отримання великої кількості молекул АТФ. Якщо трипаносома живе в організмі теплокровних ссавців, їй досить АТФ, що утворюється в процесі гліколізу.
Вперше виявлене в мітохондріях тріпаносоми редагування РНК широко поширене в хлоропластах і мітохондріях вищих рослин. Знайдено воно і в соматичних клітинах ссавців, наприклад, в кишковому епітелії людини редагується мРНК гена аполіпопротеїну.
Найбільший сюрприз вчений мітохондрії піднеслі в 1979 р До того часу вважаю, что генетичний код універсальний и одні и ті ж триплети кодують однакові амінокислоти у бактерій, вірусів, грібів, рослин и тварин. Англійський дослідник Беррел зіставів структуру одного з мітохондріальніх генів теляті з послідовністю амінокіслот в кодируемой ЦІМ геном субодініці цитохромоксидази. Виявило, что генетичний код мітохондрій Великої рогатої худоби (як и людини) не просто відрізняється від універсального, ВІН "ідеальний", тобто підпорядковується Наступний правилом: "якщо два кодони ма ють два однаково нуклеотиду, а треті нуклеотиди належати до одного класу (пуринових - А , Г, або пірімідіновіх - У, Ц), то смороду кодують одну и ту ж амінокіслоту ". В універсальному коді є два винятки з цього правила: триплет АУА кодує изолейцин, а кодон АУГ - метіонін, в той час як в ідеальному коді мітохондрій обидва ці триплета кодують метіонін; триплет УГГ кодує лише триптофан, а триплет УГА - стоп-кодон. В універсальному коді обидва відхилення стосуються принципових моментів синтезу білка: кодон АУГ - ініціює, а стоп-кодон УГА зупиняє синтез поліпептиду. Ідеальний код притаманний не всім описаним мітохондрій, але ні в одній з них немає універсального коду. Можна сказати, що мітохондрії говорять на різних мовах, але ніколи - на мові ядра.
Як вже говорилося, в мітохондріальному геномі хребетних є 22 гена тРНК. Яким же чином такий неповний набір обслуговує всі 60 кодонів для амінокислот (в ідеальному коді з 64 кодонів чотири стоп-кодону, в універсальному - три)? Справа в тому, що при синтезі білка в мітохондріях спрощені кодон-антикодон взаємодії - для впізнавання використовується два з трьох нуклеотидів антикодону. Таким чином, одна тРНК дізнається все чотири представники кодонового сімейства, що відрізняються тільки третім нуклеотидів. Наприклад, лейцінових тРНК з антикодоном ГАУ встає на рибосомі навпаки кодонів ЦУУ, ЦУЦ, ЦУА і ЦУГ, забезпечуючи безпомилкове включення лейцину в поліпептидний ланцюг. Два інших лейцінових кодону УУА і УУГ впізнаються тРНК з антикодоном ААУ. В цілому, вісім різних молекул тРНК дізнаються вісім родин по чотири кодону в кожному, і 14 тРНК дізнаються різні пари кодонів, кожна з яких шифрує одну амінокислоту.
Важливо, що ферменти аміноацил-тРНК-синтетази, відповідальні за приєднання амінокислот до відповідних тРНК мітохондрій, кодуються в ядрі клітини і синтезуються на рибосомах ендоплазматичної мережі. Таким чином, у хребетних тварин все білкові компоненти мітохондріального синтезу поліпептидів зашифровані в ядрі. При цьому синтез білків в мітохондріях подавляется циклогексимідом, блокуючим роботу евкаріотіческіх рибосом, але чутливий до антибіотиків еритроміцину і хлорамфеніколу, ингибирующим білковий синтез в бактеріях. Цей факт служить одним з аргументів на користь походження мітохондрій з аеробних бактерій при симбиотическом освіту евкаріотіческіх клітин [ 4 ].
Симбиотическая теорія походження мітохондрій
Гіпотезу про походження мітохондрій і рослинних пластид з внутрішньоклітинних бактерій-ендосімбіонтов висловив Р.Альтман ще в 1890 р За століття бурхливого розвитку біохімії, цитології, генетики і що з'явилася півстоліття тому молекулярної біології гіпотеза переросла в теорію, засновану на великому фактичному матеріалі. Суть її така: з появою фотосинтезуючих бактерій в атмосфері Землі накопичувався кисень - побічний продукт їх метаболізму. З ростом його концентрації ускладнювалася життя анаеробних гетеротрофов, і частина з них для отримання енергії перейшла від безкисневого бродіння до окислювального фосфорилювання. Такі аеробні гетеротрофи могли з більшим ККД, ніж анаеробні бактерії, розщеплювати органічні речовини, які утворюються в результаті фотосинтезу. Частина вільно живуть аеробів була захоплена анаеробами, але не "переварена", а збережена як енергетичних станцій, мітохондрій. Не варто розглядати мітохондрії як рабів, захоплених у полон, щоб постачати молекулами АТФ нездатні до дихання клітини. Вони швидше "істоти", ще в протерозої що знайшли для себе і свого потомства найкраще з притулків, де можна витрачати найменші зусилля, не піддаючись ризику бути з'їденими.
На користь симбіотичної теорії свідчать численні факти:
- збігаються розміри і форми мітохондрій і вільно живуть аеробних бактерій; ті і інші містять кільцеві молекули ДНК, не пов'язані з гистонами (на відміну від лінійних ядерних ДНК);
- по нуклеотидних послідовностей рибосомні і транспортні РНК мітохондрій відрізняються від ядерних, демонструючи при цьому дивовижну схожість з аналогічними молекулами деяких аеробних грамнегативних еубактерій;
- мітохондріальні РНК-полімерази, хоча і кодуються в ядрі клітини, відзначено зниження рифампіцином, як і бактеріальні, а евкаріотіческіе РНК-полімерази нечутливі до цього антибіотика;
- білковий синтез в мітохондріях і бактеріях пригнічується одними і тими ж антибіотиками, що не впливають на рибосоми евкаріот;
- ліпідний склад внутрішньої мембрани мітохондрій і бактеріальної плазмалемми схожий, але сильно відрізняється від такого зовнішньої мембрани мітохондрій, гомологичной іншим мембран евкаріотіческіх клітин;
- Крісті, утворені внутрішньої мітохондріальної мембраною, є еволюційними аналогами мезосомних мембран багатьох прокаріотів;
- до сих пір збереглися організми, що імітують проміжні форми на шляху до утворення мітохондрій з бактерій (примітивна амеба Pelomyxa не має мітохондрій, але завжди містить ендосімбіотіческіе бактерії).
Існує уявлення, що різні царства евкаріот мали різних предків і ендосимбіоз бактерій виникав на різних етапах еволюції живих організмів. Про це ж говорять відмінності в будові мітохондріальних геномів найпростіших, грибів, рослин і вищих тварин. Але у всіх випадках основна частина генів з промітохондрій потрапила в ядро, можливо, за допомогою мобільних генетичних елементів. При включенні частини геному одного з симбіонтів в геном іншого інтеграція симбіонтів стає незворотною.
Новий геном може створювати метаболічні шляхи, що призводять до утворення корисних продуктів, які не можуть бути синтезовані жодним з партнерів окремо. Так, синтез стероїдних гормонів клітинами кори надниркових залоз є складний ланцюг реакцій, частина яких відбувається в мітохондріях, а частина - в ендоплазматичної мережі [ 5 ]. Захопивши гени промітохондрій, ядро отримало можливість надійно контролювати функції симбіонту. У ядрі кодуються всі білки і синтез ліпідів зовнішньої мембрани мітохондрій, більшість білків матриксу і внутрішньої мембрани органел. Найголовніше, що ядро кодує ферменти реплікації, транскрипції і трансляції мтДНК, контролюючи тим самим зростання і розмноження мітохондрій. Швидкість зростання партнерів по симбіозу повинна бути приблизно однаковою. Якщо господар буде рости швидше, то з кожним його поколінням число симбіонтів, що припадають на одну особину, зменшуватиметься, і, врешті-решт, з'являться нащадки, які не мають мітохондрій. Ми знаємо, що в кожній клітині організму, розмножується статевим шляхом, міститься багато мітохондрій, реплицирующихся свої ДНК в проміжку між поділами хазяїна. Це є гарантією того, що кожна з дочірніх клітин отримає принаймні одну копію геному мітохондрії.
цитоплазматична спадковість
Крім кодування ключових компонентів дихального ланцюга і власного білоксинтезуючого апарату, мітохондріальний геном в окремих випадках бере участь у формуванні деяких морфологічних і фізіологічних ознак. До таких ознак належать характерні для ряду видів вищих рослин синдром NCS (non-chromosomal stripe, нехромосомной кодується плямистість листя) і цитоплазматична чоловіча стерильність (ЦМС), що призводить до порушення нормального розвитку пилку. Прояв обох ознак обумовлено змінами в структурі мтДНК. При ЦМС спостерігаються перебудови геномів мітохондрій в результаті рекомбінаційних подій, що ведуть до ділок, дуплікації, інверсія або Інсерція певних нуклеотидних послідовностей або цілих генів. Такі зміни можуть викликати не тільки пошкодження наявних генів, але і поява нових працюючих генів.
Цитоплазматична спадковість, на відміну від ядерної, не підкоряється законам Менделя. Це пов'язано з тим, що у вищих тварин і рослин гамети від різних статей містять несумісні кількості мітохондрій. Так, в яйцеклітині миші є 90 тис. Мітохондрій, а в сперматозоїді - лише чотири. Очевидно, що в заплідненої яйцеклітини мітохондрії переважно або тільки від жіночої особини, тобто успадкування всіх мітохондріальних генів материнське. Генетичний аналіз цитоплазматичної спадковості утруднений через ядерно-цитоплазматичних взаємодій. У разі цитоплазматичної чоловічої стерильності мутантний мітохондріальний геном взаємодіє з певними генами ядра, рецесивні аллели яких необхідні для розвитку ознаки. Домінантні аллели цих генів як в гомо-, так і в гетерозиготному стані відновлюють фертильність рослин незалежно від стану мітохондріального геному.
Вивчення геномів мітохондрій, їх еволюції, що йде за специфічними законами популяційної генетики, взаємин між ядерними і мітохондріальними генетичними системами, необхідно для розуміння складної ієрархічної організації евкаріотіческой клітини і організму в цілому.
З певними мутаціями в мітохондріальної ДНК або в ядерних генах, що контролюють роботу мітохондрій, пов'язують деякі спадкові хвороби і старіння людини [ 6 ]. Накопичуються дані про участь дефектів мтДНК в канцерогенезі. Отже, мітохондрії можуть бути мішенню хіміотерапії раку. Є факти про тісну взаємодію ядерного і мітохондріального геномів в розвитку ряду патологій людини. Множинні делеции мтДНК виявлені у хворих з важкою м'язовою слабкістю, атаксією, глухотою, розумовою відсталістю, наследующихся по аутосомно-домінантним типом. Встановлено статевої диморфізм в клінічних проявах ішемічної хвороби серця, що швидше за все обумовлено материнським ефектом - цитоплазматичної спадковістю. Розвиток генної терапії вселяє надію на виправлення дефектів в геномах мітохондрій в доступному для огляду майбутньому.
Робота виконан за ПІДТРИМКИ Російського фонду фундаментальних ДОСЛІДЖЕНЬ. Проект 01-04-48971.
Автор вдячний аспіранту М.К.Іванову, створив малюнки до статті.
література
1. Янковський М.К., Боринская С.А. Наша історія, записана в ДНК // Природа. 2001. №6. С.10-18.
2. Мінченко А.Г., Дударєва Н.А. Мітохондріальний геном. Новосибірськ, 1990..
3. Гвоздьов В.А. // Сорос. образоват. журн. 1999. №10. С.11-17.
4. Маргеліс Л. Роль симбіозу в еволюції клітини. М., 1983.
5. Скулачев В.П. // Сорос. образоват. журн. 1998. №8. С.2-7.
6. Ігамбердіев А.У. // Сорос. образоват. журн. 2000. №1. С.32-36.
Що ж представляє із себе геном мітохондрій, що відрізняється у людини і інших тварин від такого у рослин, грибів і найпростіших і за розміром, і за формою, і за генетичною ємності?
Як працює і як виник мітохондріальний геном у різних таксонів?
Яким же чином такий неповний набір обслуговує всі 60 кодонів для амінокислот (в ідеальному коді з 64 кодонів чотири стоп-кодону, в універсальному - три)?